
|
|
VII. 2. Конструктивный метаболизмКонструктивный метаболизм (пластический обмен) – совокупность биосинтетических реакций. Для биосинтеза основных клеточных компонентов микроорганизмы используют соединения-предшественники. Если эти предшественники находятся в окружающей среде, они непосредственно вовлекаются в различные биосинтетические пути, если же их нет, то они синтезируются из доступных исходных продуктов. Биосинтез аминокислот и белков. Большинство микроорганизмов способны синтезировать все необходимые им аминокислоты. Основными исходными соединениями для синтеза аминокислот являются пируват (образуется в гликолитическом цикле), α-кетоглутарат и фумарат (образуются в ЦТК). Азот вводится в аминокислоты посредством реакций переаминорования, только L-аланин, L-глутамат и аспартат образуются путем прямого аминирования. Аминокислоты идут на биосинтез белков клетки. Микроорганизмы способны синтезировать несколько тысяч различных белков. Информация об этих белках закодирована в последовательности нуклеотидов ДНК. Синтез белков осуществляется в цитоплазме на рибосомах. Биосинтез углеводов. Фототрофные организмы образуют гексозы в результате восстановления СО2. Гексозы трансформируются в крахмал, целлюлозу и другие полисахариды. В клетках других организмов углеводы образуются из неуглеводных предшественников (аминокислот, глицерина, молочной кислоты) в процессе глюконеогенеза. Биосинтез липидов. В клетках эубактерий компонентами липидов являются в основном насыщенные жирные кислоты, полиненасыщенные найдены только у цианобактерий. Жирные кислоты синтезируются отдельно, а затем с помощью эфирной связи включаются в липиды. Биосинтез нуклеотидов и нуклеиновых кислот. Большинство микроорганизмов способны к синтезу пуриновых и пиримидиновых оснований de novo из низкомолекулярных соединений. Пуриновые нуклеотиды построены на основе фосфорибозилпирофосфата, а пиримидиновые формируются в серии последовательных превращений, начиная с карбомоилфосфата. Синтезированные нуклеотиды идут на синтез нуклеиновых кислот, коферментов, богатых энергией соединений. Контрольные вопросы и задания 1. Дайте определение метаболизма. 2. Какие способы получения энергии известны у микроорганизмов? 3. Что такое брожение? Какие виды брожения вам известны? 4. Какие продукты образуются при гомо- и гетероферментативном брожении? 5. Назовите ключевой фермент спиртового брожения. 6. Охарактеризуйте пропионовокислое брожение. 7. Какие бактерии осуществляют маслянокислое брожение? 8. Какой процесс называют дыханием? Какие виды дыхания вам известны? 9. Какие бактерии способны к фотосинтезу? 10. Охарактеризуйте строение фотосинтетического аппарата у фотосинтезирующих бактерий. 11. Какими путями происходит фиксация СО2 у бактерий? Список рекомендуемой литературы 1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия: Учебник.– М.: Медицина, 1998.– 704 с. 2. Градова Н.Б., Бабусенко Е.С., Горнова И.Б., Гусарова Н.А. Лабораторный практикум по общей микробиологии. – М.: ДеЛи Принт, 2001. – 131 с. 3. Гусев М.В. Микробиология: Учебник для студ. биол. специальностей вузов /М.В. Гусев, Л.А.Минеева. – М.: Изд. центр «Академия», 2003. – 464 с. 4. Емцев В.Т., Мишустин Е.Н. Микробиология: Учебник для вузов. – М.: Дрофа, 2005. – 445 с. 5. Коротяев А.И., Бабичев С.А. Медицинская микробиология, иммунология и вирусология: Учебник для вузов. – СПб.: СпецЛит, 2002. – 591 с. 6. Поздеев О.К. Медицинская микробиология / Под ред. акад. РАМН В.И. Покровского. – М.:ГЭОТАР-МЕД, 2001. – 768 с. 7. Практикум по микробиологии: Учеб. пособие для студ. высших учебных заведений / А.И. Нетрусов, М.А. Егорова и др.; Под ред. А.И. Нетрусова. – М: Изд. центр «Академия», 2005. – 608 с. 8. Справочник биохимика.– Пер. с англ. / Досон Р., Эллиот Д., Эллиот У., Джонс К. – М.: Мир, 1991.– 544 с. 9. Шлегель Г. Общая микробиология. – М.: Мир, 1987. – 556 с.
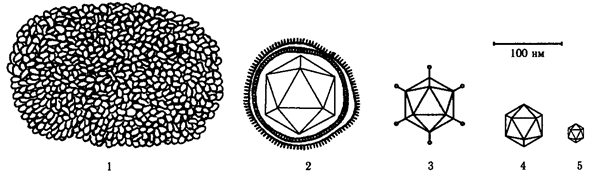
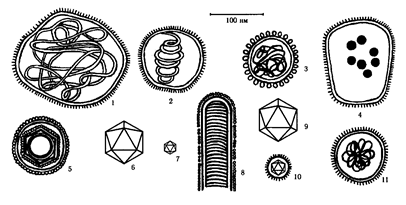
Глава VIII. ВИРУСЫ Вирусы (от лат. virus – яд) – это внеклеточная форма жизни, обладающая собственным геномом и способная к воспроизведению только в клетках живых организмов. Вирусы проникают в растительные, животные ткани, бактерии (вирусы бактерий называют бактериофагами). Вирусы являются внутриклеточными паразитами на генетическом уровне и используют для своего размножения белоксинтезирующий аппарат клетки-хозяина. Вирусы классифицируют на основе их морфологических, химических, биофизических свойств и особенностей репродукции. Виды объединяют в роды и семейства. Название всех вирусных родов оканчивается словом «virus» (Enterovirus, Reovirus), для названия семейств используется суффикс «idae» (Poxviridae, Herpesviridae), а подсемейств «inae» (Chordopoxvirinae, Entomopoxvirinae). VIII.1. Строение вирусов Все вирусы условно делятся на простые и сложные. Простые вирусы («голые») состоят из одной или нескольких молекул ДНК (ДНК-содержащие вирусы) или РНК (РНК-содержащие вирусы), заключенных в белковую оболочку – капсид (от греч. capsa – ящик). Сложные вирусы («одетые») имеют вторую оболочку – суперкапсид, который наряду с белками содержит липопротеидную мембрану, углеводы и неструктурные белки – ферменты. Суперкапсидные вирусные белки могут образовывать различные шипы и обладают жизненноважными для вирусов функциями: распознают клеточные рецепторы, связываются с ними, обеспечивают проникновение вирусов и их распространение в организме, обладают антигенными свойствами. Формы вирусов многообразны: палочковидные, нитевидные, сферические, кубовидные, булавовидные и др. (рис. 38; 39).
Рис.38. Формы и относительные размеры ДНК-содержащих вирусов животных 1 – Poxviridae; 2 – Herpesviridae; 3 – Adenoviridae; 4 – Papovaviridae; 5 – Parvoviridae
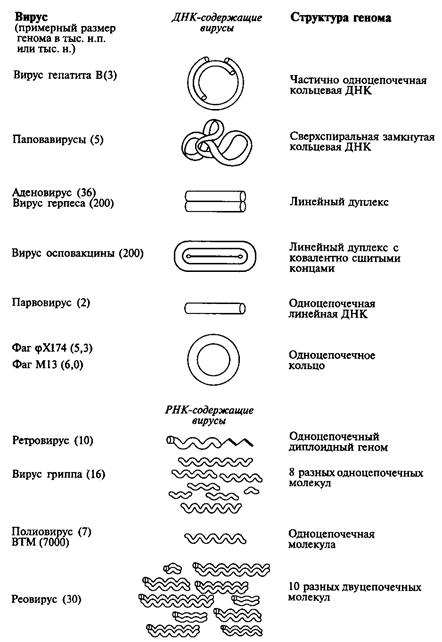
ДНК или РНК вирусов могут быть одно- и двуцепочечными, кольцевыми и линейными (рис. 40). Значения масс ДНК вирусов находятся в пределах 1·106 – 200·106; РНК – 1·106 – 15·106 дальтона.
Рис. 39. Формы и относительные размеры РНК-содержащих вирусов животных 1 – Paramyxoviridae; 2 – Orthomyxoviridae; 3 – Coronaviridae; 4 – Arenaviridae; 5 – Retroviridae;
Рис.40. Структура геномов вирусов VIII.2. Репродукция вирусов Репликация генетического материала вирусов, а также экспрессия вирусных генов осуществляется при помощи механизмов репликации, транскрипции и трансляции инфицированной клетки-хозяина.
Рис. 41. Проникновение вируса У вирусов, имеющих липидную мембрану, есть второй путь проникновения в клетку. При этом способе происходит слияние двух мембран (вирусной и клеточной), с образованием сквозного отверстия, через которое содержимое вируса входит в клетку. Этот путь использует вирус СПИДа. Вирусы бактерий (бактериофаги) проникают в клетку иным способом. Так, бактериофаг l (рис. 42) прикрепляется к клеточной стенке нитевидным хвостом и с его помощью вводит ДНК в бактериальную клетку. Репродуктивный цикл вируса начинается после высвобождения его генома в цитоплазму. Для образования (сборки) дочерних вирусных частиц необходим синтез генетического вируса, экспрессия генетического материала и образование вирусных белков. В зависимости от типа генетического материала (ДНК и РНК), образование дочерних копий геномов протекает по-разному. РНК-содержащие вирусы делятся на три группы: 1) вирусы, содержащие однонитевой геном с положительной полярностью (+РНК-цепь), т.е. с нуклеотидной последовательностью, соответствующей таковой у мРНК; 2) вирусы, содержащие однонитевой геном с негативной полярностью (-РНК-цепь); 3) вирусы, имеющие двунетевые геномы. Если (+) РНК-цепь попадает в клетку, то ее белоксинтезирующий аппарат может немедленно транслировать РНК, производя вирусные белки, в том числе ферменты, необходимые для репродукции вируса. (-) РНК-цепь не может выполнять функции мРНК, поэтому вирус внедряет в клетку не только геном, но и фермент, умеющий снимать с этого генома комплементарные копии. Фермент (РНК-зависимая РНК-полимераза) копирует вирусный геном, образуя (+)РНК, которая выступает в качестве матрицы для синтеза вирусных белков. Двунетевые геномы вирусов сегментированы (т.е. состоят из нескольких разных молекул), их размножение происходит по варианту, близкому к предыдущему. Имеется один класс вирусов, содержащих (+)РНК-цепь – ретровирусы. В эту группу входят вирус иммунодефицита человека (ВИЧ) и некоторые возбудители злокачественных новообразований. В вирусном геноме закодирован необычный фермент (обратная транскриптаза, или ревертаза). Этот фермент попадает в заражаемую клетку вместе с вирусной РНК и обеспечивает синтез ее ДНК-копии сначала в одноцепочечной форме, затем в двуцепочечной ДНК, которая встраивается в хромосому клетки-хозяина. Провирусные гены (гены вируса в хромосомах хозяина) транскрибируются в ядре клетки в (+)РНК-транскрипты. Одни из них становятся геномом нового потомства вирусов, а другие используются для трансляции белков, необходимых для сборки вирусных частиц. Репликация ДНК-содержащих вирусов (двуцепочечных) осуществляется по схеме ДНК ® РНК ® белок. Молекулы РНК образуются в результате транскрипции вирусных ДНК в клеточном ядре хозяйским ферментом ДНК – зависимой РНК-полимеразой. Транскрибируется только одна из нитей вирусной ДНК. Синтез ДНК на РНК-матрицы происходит в результате реакций, катализируемой обратной транскриптазой; сначала синтезируется (-) нить ДНК, а затем на ней строится (+) нить. Если вирусы содержат одноцепочечную ДНК (+ или -) в ходе репликации с использованием фермента клетки-хозяина образуется двуцепочечная ДНК, с которой транскрибируется мРНК. Конечной стадией репродуктивного цикла является высвобождение дочерних вирионов. Выход вирусов из клетки может сопровождаться гибелью клетки-хозяина (литический путь). Фермент лизоцим, кодируемый вирусным геном на поздних стадиях развития, разрушает клеточную стенку и вызывает лизис клетки. Ретровирусные частицы покидают клетку, не убивая ее. Включенный в состав клеточной хромосомы вирусный геном, передается дочерним клеткам, свойства которых при этом могут меняться. Альтернативным литическому пути является лизогенный путь. Размножение вирусов в данном случае блокируется. Вирусная хромосома включается в геном клетки-хозяина и затем размножается вместе с ним, либо образует плазмиду. Это может привести к генетическим изменениям, в результате которых начинается неконтролируемый рост клеток и превращение их в раковые. Контрольные вопросы и задания 1. Что представляют собой вирусы? 2. Какие организмы способны поражать вирусы? 3. Как происходит проникновение вирусов в клетки? 4. Расскажите о репликации вирусного генома. 5. Что происходит с клеткой-хозяина при литическом и лизогенном способах размножения вирусов? Список рекомендуемой литературы 1. Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и применение. – Пер. с англ. – М.: Мир, 2002.– 589 с. 2. Грин Н., Статут У., Тейлор Д. Биология: В 3 т. – Т. 1. – Пер с англ. / Под ред. Р. Сопера. – М.: Мир, 1996. – 368 с. 3. Гусев М.В. Микробиология: Учебник для студ. биол. специальностей вузов /М.В. Гусев, Л.А.Минеева. – М.: Изд. центр «Академия», 2003. – 464 с. 4. Жимулев И.Ф. Общая и молекулярная генетика: Учеб. пособие. – Новосибирск: Изд-во Новосиб. ун-та: Изд-во Сиб. ун-та, 2002.– 459 с. 5. Емцев В.Т., Мишустин Е.Н. Микробиология: Учебник для вузов. – М.: Дрофа, 2005. – 445 с. 6. Коничев А.С. Молекулярная биология. – М.: Изд. центр «Академия», 2003.– 400 с. 7. Коротяев А.И., Бабичев С.А. Медицинская микробиология, иммунология и вирусология: Учебник для вузов. – СПб.: СпецЛит, 2002. – 591 с. 8. Поздеев О.К. Медицинская микробиология / Под ред. акад. РАМН В.И. Покровского. – М.:ГЭОТАР-МЕД, 2001. – 768 с. 9. Практикум по микробиологии: Учеб. пособие для студ. высших учебных заведений / А.И. Нетрусов, М.А. Егорова и др.; Под ред. А.И. Нетрусова. – М: Изд. центр «Академия», 2005. – 608 с. 10. Чемерилова В.И. Основы генной инженерии. – Иркутск: Изд-во Иркутского университета, 1998. –140 с. 11. Ченцов Ю.С. Введение в клеточную биологию.– М.: Изд. центр «Академия», 2004.– 495 с. 12. Шлегель Г. Общая микробиология. – М.: Мир, 1987. – 556 с. 13. Эллиот В. Биохимия и молекулярная биология / В.Эллиот, Д. Эллиот; Под ред. А.И. Арчакова, М.П. Кирпичникова, А.Е. Медведева, В.П. Скулачева. – Пер. с англ. О.В. Добрыниной, И.С. Севериной, Е.Д. Скоцеляс и др. – М.: МАИК «Наука/ ИНтерпериодика», 2002. – 446 с.
Глава IX. Рост и распространение микроорганизмов в природе определяется условиями внешней среды. На жизнедеятельность микроорганизмов оказывают влияние физические и химические факторы. VIII. 1. Физические факторы Среди физических факторов наибольшее значение имеют температура, влажность, кислотность среды, наличие кислорода, влияние лучистой энергии, давления. Температура. По отношению к температуре микроорганизмы делятся на три группы: психофилы, мезофиллы и термофилы. Психофильные виды (от греч. psychros – холод + phileo – любить) растут в диапазоне – 10 до + 200С. Среди них есть как облигатные так и факультативные виды. Облигатные психофилы приспособились к устойчивым холодным условиям (глубины морей и океанов, ледяные пещеры, высокогорные районы), облигатные психофилы обитают в неустойчивых холодных условиях. Мезофильные виды (от греч. mesos – тепло + phileo – любить) лучше растут в пределах 20 – 400С. В эту группу входят большинство микроорганизмов, среди них много патогенных и условно-патогенных. Термофильные виды (от греч. thermo – тепло + phileo – любить) растут при температуре 400С и выше. Среди термофилов выделяют термотолерантные факультативные, облигатные и экстремальные виды. Термотолерантные виды растут в пределах от 10 до 55–600С, оптимум лежит при 35–40оС. Факультативные термофилы имеют максимальную температуру роста между 50 и 65оС, но способны размножаться и при комнатной температуре. При температуре около 70оС растут облагатные термофилы, их рост прекращается при 40оС. Температурный оптимум для экстремальных термофилов лежит в пределах 80–105оС. К экстремальным термофилам относятся организмы из группы архебактерий. Основное место их обитания – горячие источники. Механизмы, обеспечивающие существование архебактерий при повышенных температурах до конца не известны. Предполагают, что определенную роль в этом играют липидные компоненты клеточных мембран с высоким содержанием длинноцепочечных С17–С19 насыщенных жирных кислот с разветвленными цепями, а также высокая термостабильность белков, ферментов и структурных компонентов клетки. Высокая температура вызывает коагуляцию структурных белков и ферментов микроорганизмов. Большинство вегетативных форм гибнет при температуре 60оС в течение 30 мин, а при 80–100оС через мин. Температурные воздействия применяют для стерилизации – полного удаления микроорганизмов из различных сред и обеззараживания предметов. Существует несколько способов стерилизации. Самыми простыми являются прокаливание и кипячение. Эффективным методом является автоклавирование – обработка горячим паром под высоким давлением. Стерилизация сухим жаром проводится в сухожаровых шкафах при 160оС в течение 2 ч, что позволяет уничтожать не только вегетирующие клетки, но и споры бактерий. В пищевой промышленности используют пастеризацию (нагревание до 60–80оС в течение 10–30 мин). Кислотность среды. Для большинства микроорганизмов оптимальные значения кислотности среды около рН 7 (нейтрофилы), при крайних значениях рН1 (ацидофилы) и рН 11(алкалофилы) могут существовать лишь немногие из них. Нейтрофилы развиваются в диапазоне рН 4–9 (Bacillus subtilis, Streptococcus faecalis, Escherichia coli и др.). Среди нейтрофильных бактерий есть микроорганизмы, обладающие кислото- и щелочеустойчивостью (толерантностью). Для алкалофилов предпочтительна щелочная реакция среды (рН 10 и выше). Некоторые нитрат- и сульфатвосстанавливающие бактерии могут существовать при рН выше 11. Ацидофилы растут при рН ниже 6. Архебактерии Sulfolobus acidocaldarius растут при рН от 1до 5, 8, оптимальная область рН 2–3. Наличие кислорода. По отношению к молекулярному кислороду все микроорганизмы делятся на аэробы (для их роста необходим кислород) и анаэробы (кислород для роста не нужен). Среди как аэробов, так и анаэробов есть облигатные и факультативные виды. Существуют облигатно аэробные (строгие) прокариоты, которые потребляют кислород, но хорошо растут при содержании его в меньшей концентрации, чем в атмосфере. Такие микроорганизмы называют микроарофильными. Среди облигатных аэробов существуют различия в устойчивости к высоким уровням О2 в среде. 100% молекулярный кислород подавляет рост всех облигатных аэробов. Многие из облигатных анаэробов не выносят присутствия даже незначительных количеств молекулярного кислорода в среде и быстро погибают. К числу строгих анаэробов относятся представители родов Bacteroides, Fusobacterium, Butyrivibrio и др. Есть виды умеренно (Clostridium tetani, C. carnis, C. tertium) или достаточно высоко (C. perfringens, C. acetobutylicum) толерантные к О2. Молочнокислые бактерии относятся к аэротолерантным анаэробам, они обладают метаболизмом анаробного типа, но могут расти и в присутствии воздуха. Известны микроорганизмы, которые приспособились, в зависимости от наличия или отсутствия О2 в среде, переключаться с одного метаболитического пути на другой, например, с дыхания на брожение, и наоборот. Такие организмы называют факультативными анаэробами или факультативными аэробами. К ним относятся, например, энтеробактерии. Излучение. Свет является необходимым фактором для фотосинтезирующих микроорганизмов, но для большинства других бактерий свет губителен. Спектр солнечной активности содержит неионизирующие (УФ- и инфракрасные лучи) и ионизирующее (например, g-лучи). Наибольший микробицидный эффект оказывают коротковолновые УФ-лучи (250–270 нм), которые действуют на нуклеиновые кислоты. Повреждения ДНК ведут к появлению нежизнеспособных мутантов. Влажность. Микроорганизмам для роста и размножения необходима влага. Жизнедеятельность большинства бактерий прекращается при относительной влажности среды ниже 30%. Время отмирания бактерий при высушивании различно, например, холерный вибрион погибает за 2 суток, микобактерии – за 90 суток. Метод искусственного высушивания (лиофилизацию) используют для сохранения имуннобиологических препаратов (вакцин, сывороток), а также для консервирования и длительного сохранения микроорганизмов. Высушивание применяют при консервировании сухих продуктов и изготовлении сухих концентратов пищевых продуктов. VIII. 2. Химические факторы Химические вещества могут подавлять рост и размножение микроорганизмов. Действие веществ зависит от их концентрации, природы, особенностей микроорганизма, факторов внешней среды. В медицинской практике широко используют различные виды дезинфектантов и антисептиков. Дезинфектанты используют для обработки помещений и предметов, антисептики – для обработки живых тканей. Галогены и галогенсодержащие препараты (препараты йода и хлора) используют как дезинфектанты и антисептики. Они взаимодействуют с гидроксильными группами белков, нарушая их структуру. Альдегиды (формальдегид, глутаральдегид) алкилируют сульфгидрильные, карбоксильные и аминогруппы белков и других соединений, вызывая гибель микроорганизмов. Кислоты (борная, бензойная, уксусная,салициловая), щелочи (раствор аммиака ), металлы (нитрат серебра, сульфат меди, хромат ртути), красители (бриллиантовый зеленый, метиленовый синий), окислители (перекись водорода, перманганат калия) применяют как антисептики. Фенолы (гексахлоран, хлорофен, тимол)денатурируют белки и нарушают структуру клеточной стенки, широко используются в качестве дезинфектантов. В отличие от дезинфектантов и антисептиков, имеющих неспецифическое действие, химиотерапевтические средства проявляют избирательное противомикробное действие. Антибактериальные, противогрибковые, антипротозойные препараты тормозят рост, либо вызывают гибель микроорганизмов. Антибиотики(от греч. anti – против + bios – жизнь) – широкий класс антибактериальных препаратов, имеют различный механизм действия. Они способны подавлять процессы синтеза компонентов клеточной стенки, синтеза белка и нуклеиновых кислот и др. Механизм действия большинства противогрибковых средств связан с нарушением синтеза стеролов, входящих в состав клеточной стенки. Противопротозойные препараты угнетают уникальные ферменты простейших. Противовирусные препараты ингибируют репликацию вирусов. Контрольные вопросы и задания 1. Какие факторы внешней среды оказывают влияние на жизнедеятельность микроорганизмов? 2. На какие группы делятся микроорганизмы в зависимости от температурного фактора, кислотности среды? 3. Расскажите об аэробных и анаэробных микроорганизмах. 4. Какую роль играет в жизнедеятельности микроорганизмов вода? 5. Каковы механизмы действия химических факторов? Список рекомендуемой литературы 1. Градова Н.Б., Бабусенко Е.С., Горнова И.Б., Гусарова Н.А. Лабораторный практикум по общей микробиологии. – М.: ДеЛи Принт, 2001. – 131 с. 2. Емцев В.Т., Мишустин Е.Н. Микробиология: Учебник для вузов. – М.: Дрофа, 2005. – 445 с. 3. Нетрусов А.И., Бонч-Осмоловская Е.А., Горленко В.М. и др. Экология микроорганизмов. – М: Изд. центр «Академия», 2005.– 272 с. 4. Поздеев О.К. Медицинская микробиология / Под ред. акад. РАМН В.И. Покровского. – М.:ГЭОТАР-МЕД, 2001. – 768 с. 5. Практикум по микробиологии: Учебное пособие для студентов высших учебных заведений / А.И. Нетрусов, М.А. Егорова и др.; Под ред. А.И. Нетрусова. – М: Изд. центр «Академия», 2005. – 608 с. 6. Шлегель Г. Общая микробиология. – М.: Мир, 1987. – 556 с. Глава X. Вездесущность микроорганизмов обусловлена их разнообразием и способностью использовать любые источники питания. В биосфере нет такой среды, в которой не встречались бы микроорганизмы. Образуя сложные ассоциации (микробные сообщества), они играют колоссальную роль в природе, круговороте веществ, поддерживают динамическое равновесие биосферы.   Система охраняемых территорий в США Изучение особо охраняемых природных территорий(ООПТ) США представляет особый интерес по многим причинам...  Что делает отдел по эксплуатации и сопровождению ИС? Отвечает за сохранность данных (расписания копирования, копирование и пр.)...  Что вызывает тренды на фондовых и товарных рынках Объяснение теории грузового поезда Первые 17 лет моих рыночных исследований сводились к попыткам вычислить, когда этот...  ЧТО ПРОИСХОДИТ, КОГДА МЫ ССОРИМСЯ Не понимая различий, существующих между мужчинами и женщинами, очень легко довести дело до ссоры... Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
|
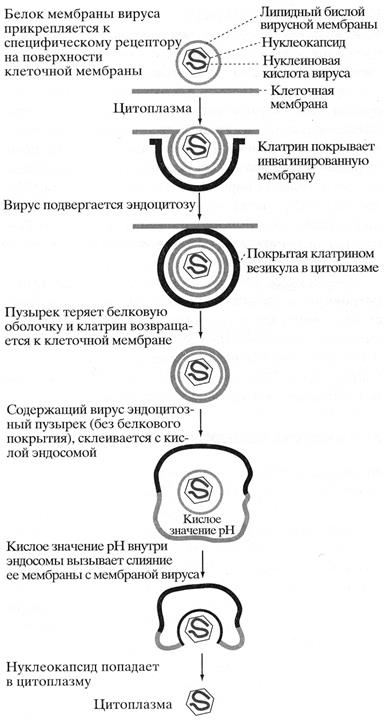 Живая клетка окружена липидной бислойной мембраной, которая препятствует проникновению вируса. Однако вирусы имеют способы преодоления этих барьеров. Некоторые вирусы используют для проникновения в клетку рецептор-опосредуемый эндоцитоз. На первом этапе вирион (внеклеточная форма вируса) связывается с поверхностью клетки при помощи покровного белка, комплементарного специфическому рецептору, расположенному снаружи клетки-хозяина. После прикрепления комплекс вирус-рецептор движется в мембране к углублению – окаймленной ямке, внутренняя поверхность которой выстлана белком клатрином. Ямка впячивается до поглощения вириона внутрь клетки. Клатрин возвращается к клеточной мембране, а содержащий вирус пузырек сливается с цитоплазматическим пузырьком – эндосомой. Кислая среда внутри эндосомы вызывает дисоциацию вируса от мембранного рецептора клетки – хозяина. В случае, если вирус имеет мембранную оболочку, она сливается с мембраной эндосомы – содержимое вируса оказывается в цитоплазме. Здесь вирус теряет белковую оболочку, высвобождая в цитоплазму генетический материал (рис. 41). Механизм проникновения вируса, лишенного мембраны, из эндосомы в цитоплазму не выяснен.
Живая клетка окружена липидной бислойной мембраной, которая препятствует проникновению вируса. Однако вирусы имеют способы преодоления этих барьеров. Некоторые вирусы используют для проникновения в клетку рецептор-опосредуемый эндоцитоз. На первом этапе вирион (внеклеточная форма вируса) связывается с поверхностью клетки при помощи покровного белка, комплементарного специфическому рецептору, расположенному снаружи клетки-хозяина. После прикрепления комплекс вирус-рецептор движется в мембране к углублению – окаймленной ямке, внутренняя поверхность которой выстлана белком клатрином. Ямка впячивается до поглощения вириона внутрь клетки. Клатрин возвращается к клеточной мембране, а содержащий вирус пузырек сливается с цитоплазматическим пузырьком – эндосомой. Кислая среда внутри эндосомы вызывает дисоциацию вируса от мембранного рецептора клетки – хозяина. В случае, если вирус имеет мембранную оболочку, она сливается с мембраной эндосомы – содержимое вируса оказывается в цитоплазме. Здесь вирус теряет белковую оболочку, высвобождая в цитоплазму генетический материал (рис. 41). Механизм проникновения вируса, лишенного мембраны, из эндосомы в цитоплазму не выяснен.